 2
2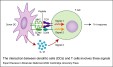 3
3 4
4 5
5 6
6 ,7a
,7a ,7b
,7b 8
8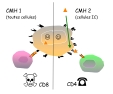 9
9 10
10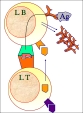 11
11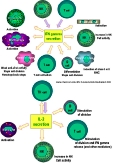 12
12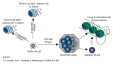 13
13 14
14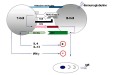 15
15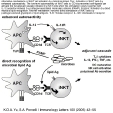 16
16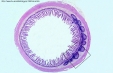
| accueil | immun 1-15: Cellules présentatrices d'antigène: apprêtement et présentation des antigènes; ; coopération cellulaire: immunogénicité, évolution de la réponse anticorps et mémorisation |
<-- cours --> |
objectifs pédagogiques (*) |
|
|
- décrire succinctement les mécanismes de présentation des antigènes endogènes et exogènes | |
définitions générales - messages |
|
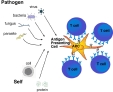
| cellule présentatrice d'antigène (CPA) | = cellule immunocompétente capable de présenter des antigènes aux lymphocytes T, sous forme de complexes associant le CMH de classe 2 et un épitope T. Les CPA, une fois activées, captent les antigènes et se déplacent des tissus vers les zones riches en lymphocytes (organes lymphoïdes secondaires, follicules disséminés dans les muqueuses). Elles produisent en même temps des cytokines qui activent les lymphocytes (IL12 et cytokines pro-inflamamatoires). |
La plupart des CPA ont besoin d'être activées pour présenter efficacement (l'activation se fait le plus souvent grâce à la reconnaissance non spécifique de signaux de danger d'origine microbienne ou tissulaire via les TLR= Toll-like receptors).(cf tableau 1) ( fig1: présentation des antigènes aux lymphocytes par les cellules dendritiques et fig2: présentation cellule dendritique-lymphocyte) (fig3: presentation des Ag dans les organes lymphoïdes II: constitution de follicules lymphoïdes II) |
|
| Cas particuliers: - les cellules dendritiques ont également la capacité à présenter des antigènes via le CMH1 à des lymphocytes T CD8 (dans ce cas, il y a activation/ différencation du LT, et non pas activité cytotoxique contre la cellule présentatrice). - la présentation des antigènes dans le thymus assure l'éducation (sélection- et +) via des mécanismes différents |
|
| apprêtement antigénique | = "antigen processing" = mécanisme réalisé dans le cytoplasme des CPA, qui produit des épitopes T (par protéolyse ménagée des antigènes) et qui associe ces épitopes T aux antigènes du CMH pour les présenter à la membrane cellulaire. - l'équipement en protéases de la CPA découpe les antigènes protéiques en épitopes séquentiels d'une dizaine d'acides aminés. Ces protéases clivent selon des séquences déterminées, qui sont peu fréquentes dans les protéines normales de la cellules (elles sont plus susceptibles de cliver des protéines étrangères). - 2 mécanismes distincts sont réalisés selon la classe de CMH (cf ci dessous). (fig4: association de l' épitope T au CMH de type 1 et 2: "niche à peptide" ) |
| épitope endogène | = épitope T issu de la dégradation d'un antigène présent dans le cytoplasme cellulaire (suite à une infection ou une parasitose intracellulaire..). Les antigènes endogènes sont découpés par des ensembles de protéases cytoplasmiques (=protéasomes), puis transloqués dans le REG où les épitopes s'associent aux CMH1. (fig5: principe de la présentation des Ag endogènes via CMH1) |
| épitope exogène | = épitope T issu de la dégradation d'un antigène capté par phagocytose ou par pinocytose. Les micro-organismes, les complexes Ag-Ac et les antigènes solubles sont dégradés par les enzymes lysosomiales dans des vacuoles, qui fusionnent ensuite avec des vacuoles d'exportation du REG à la membrane: il y a alors formation des complexes épitopes T-CMH2. (fig6: principe de la présentation des Ag exogènes via CMH2) Les macrophages ne peuvent réaliser une présentation efficace des antigènes que lorsqu'ils sont peu activés (sinon la dégradation lysosomiale est complète). |
| gamme peptidique | = ensemble des épitopes T susceptibles de s'associer à une molécule donnée du CMH, en tenant compte des contraintes stériques et chimiques qui permettent au peptide de se loger dans la "niche à peptide". Chaque molécule du CMH présente en effet des particularités stéréo-chimiques de la poche à peptide et peut de ce fait fixer ± efficacement certains épitopes. Ceci explique les variations dans la capacité de reconnaissance des antigènes en fonction du CMH. La diversité des molécules du CMH exprimée par chaque cellule lui permet de présenter un très grand nombre d'épitopes. Néanmoins, certains épitopes sont préférentiellement présentés en association avec certaines molécules du CMH : ceci explique la variation individuelle et spécifique dans les capacités de reconnaissance antigénqiue, et donc dans la résistance aux infections ou le développement de maladies auto-immunes. (fig7a: exemple d'identification de la gamme peptidique reconnue sur un antigène en association avec un CMH donné et fig7b: acides aminés reconnus par HLA-A). |
| CD4 et CD8 | = chaines protéiques constitutives du TCR de type alpha-beta, qui stabilisent la reconnaissance spécifique conjointe, en fixant respectivement le CMH2 et le CMH1. Les lymphocytes T alpha-beta matures se partagent en 2 sous-populations fonctionnelles principales, qui expriment soit le CD4, soit le CD8 (fig8: 4xII=8xI!): - les lymphocytes T CD4 sont principalement des lymphocytes producteurs de cytokines régulatrices de la réponse immune (on parle de lymphocytes T "helper"= Th, T "regulator"=TReg..) - les lymphocytes T CD8 possèdent principalement des capacités cytotoxiques ("CTL"). Ce processus a lieu durant les mécanismes de production des lymphocytes T dans le thymus (cf education). La sous-population lymphocytaire mature "double-positive" (à la fois CD4 et CD8) est généralement très minoritaire (sauf chez les porcins). |
| immunogénicité | = aptitude d'un antigène à provoquer une forte réponse immune anamnestique (antigène immunogène). L'immunogénicité d'un antigène assure en premier lieu la production d'une réponse anticorps de type secondaire (immun1-06) , ainsi qu'une immunité cellulaire (production de cytokines et éventuellement activité cytotoxique). La mémoire se constate lors de chacune des expositions ultérieures à l'antigène. La réponse anticorps secondaire s'enclenche quand la stimulation immune est suffisante pour aboutir à la différenciation des lymphocytes B, grâce aux phénomènes de coopération entre les lymphocytes T et B. Il s'agit d'interactions entre des lymphocytes qui reconnaissent les épitopes T et B d'un même antigène: les lymphocytes différenciés acquièrent des fonctions différentes des lymphocytes "naïfs" (producteurs d'IgG..). Une partie des lymphocytes acquière une durée de vie longue, assurant la mémoire immune. |
| Tous les antigènes ne sont pas immunogènes (ne pas confondre antigénicité et immunogénicité) : - les immunogènes doivent posséder à la fois des épitopes T et B pour assurer la coopération : la condition principale de l’immunogénicité est donc le caractère protéique de l’antigène, qui permet aux CPA de le fragmenter en épitopes T. - les 2 populations lymphocytaires spécifiques T et B doivent être présentes : normalement un individu ne possède pas de lymphocytes T très affins contre ses propres protéines/épitopes T (cf protection du soi). (fig9: exemple de démonstration expérimentale du rôle de l'épitope T dans l'immunogénicité) (la souris ne possède pas de lymphocyte T anti lysozyme de souris, mais possède des lymphocytes T anti lysozyme de poulet). |
|
| Antigène thymo-indépendant | cf immun1-16 |
| coopération cellulaire | = mécanisme d'interaction des lymphocytes entre eux ou avec d'autres cellules immunocompétentes, aboutissant à une modulation de leurs activités, de leurs capacités de prolifération et de leur état de différenciation. La coopération s'effectue par des échanges de signaux et de cytokines entre les cellules. (fig10: coopération entre un lymphocyte B et un lymphocyte T reconnaissant le même antigène pour la mise en place de la réponse anticorps secondaire et fig11: exemples d'effet des cytokines produites par des lymphocytes T stimulés par l'antigène) |
| mémoire immune (réponse anamnésitique) | = capacité des lymphocytes spécifiques à persister après une stimulation par un antigène, et à répondre plus rapidemment et plus intensément à une ré-exposition par le même antigène. (mémoire= capacité à stocker et réutiliser des informations= anamnèse). La mémoire est essentiellement basée sur la persistance de lymphocytes "mémoire" à durée de vie longue dans les organes lymphoïdes secondaires (une petite partie continue à circuler). Une coopération entre lymphocytes B et T est nécessaire pour obtenir des lymphocytes "mémoire" (cela ne concerne donc que des antigènes immunogènes). (fig12: différenciation mémoire d'une partie des lymphocytes B spécifiques) La stimulation des lymphocytes mémoire par l'antigène: - procure une réponse rapide (les lymphocytes mémoire s'activent et prolifèrent plus vite que les lymphocytes "naïfs") - est à la base de la réponse anticorps secondaire (réponse plus forte, plus rapide, de classe IgG et IgE/IgA). |
| commutation isotypique | = capacité d'un lymphocyte B à changer la classe d'Ig produite, sous l'effet de la coopération avec les lymphocytes T. La commutation suit un ordre au cours de la différenciation: IgM, IgG (différentes sous-classes), IgA, IgE. Une fois la différenciation fixée à un stade, la classe d'Ig produite est peu modifiable, à moins que de nouveaux mécanismes de coopération LB-LT se mettent en place. - les anticorps produits au cours de la réponse anticorps primaire sont des IgM (lymphocytes B peu ou pas différenciés, réagissant de façon autonome) - les anticorps produits au cours de la réponse anticorps secondaire sont principalement des IgG, mais aussi des IgA et/ou des IgE (lymphocytes B différenciés au cours du processus de coopération avec des lymphocytes T spécifiques du même antigène). (fig13: réponse anticorps primaire et secondaire et fig14: commutation isotypique IgE) |
schémas et figures |
|
1234 5 6,7a ,7b 89101112 13 141516
tableaux |
| tableau 1: présentation via CMH ou CD1 | CMH1 | CMH2 | Ag présentés | effets | activation de la CPA |
| quasi-toutes les cellules tissulaires (sauf cellules germinales, hématies..) | + | - (sauf anomalie) | épitopes T/CMH1 issus d'antigènes protéiques endogènes (infection intra-cellulaire, virale, bactérienne ou parasitaire..) | - reconnaissance par CTL CD8: cytolyse - destruction par NK en cas de diminution anormale de l'expression du CMH1 |
|
| nombreuses cellules immunocompétentes (cellules dendritiques..) | expression membranaire de CD1 (protéine de présentation des lipides) | un certain nombre de lipides membranaires ou microbiens | reconnaissance par lymphocytes Tgamma-delta et iNKT: production de cytokines pro-inflammatoires ou inhibitrices de l'immunité | ||
| principales cellules présentatrices (présentation à la fois d'antigènes endogènes et exogènes) | |||||
| monocyte-macrophages (et cellules apparentées) | + | + à ++ (selon état d'activation) | épitopes T/CMH2 issus de la phagocytose des bactéries, cellules, levures et parasites (et des corps étrangers) | coopération avec lymphocytes T CD4 (augmentation des capacités de phagocytose par explosion respiratoire..) | signaux de danger microbiens ou cytokines |
| lymphocytes B | + | + à ++ (selon état d'activation) | épitopes T/CMH2 isssus de l'antigène reconnu par l'anticorps | coopération avec lymphocytes T CD4 (différenciation du lymphocyte B en plasmocyte ou cellule B mémoire, commutation isotypique..) | antigène spécifique |
| cellules dendritiques | + | +++ | - épitopes T/CMH2 issus de la phagocytose des bactéries, cellules, levures et parasites - épitopes T/CMH2 issus de la pinocytose des antigènes solubles et des complexes Ag-Ac |
- coopération avec lymphocytes T CD4 (différenciation du lymphocyte T) - déplacement vers les organes lymphoïdes secondaires après captation des antigènes et activation - relargage progressif d'antigènes dans les organes lymphoïdes secondaires (stimulation des lymphocytes B). |
signaux de danger microbiens ou cytokines |
| cellules M des plaques de Peyer (et cellules apparentées) | + | + | épitopes T/CMH2 issus des antigènes présents dans le tube digestif (principalement microbiens) | coopération avec lymphocytes TCD4 et Tgamma-delta des plaques de Peyer | |
tableau 2: exemples de coopération |
conditions | effets |
| lymphocyte T CD4 et lymphocyte B | lymphocytes T et B spécifiques d'un même antigène (épitope B et T); reconnaissance de l'épitope T par le LB via CMH2 au LTh (= lymphocyte T "helper" CD4) | - sécrétion de cytokines par le lymphocyte T (les effets sur le LB varient selon les cytokines produites, c’est à dire selon le sous-type de lymphocyte Th1 ou Th2..). Cf cours régulation - différenciation du lymphocyte T (mémoire..) - différenciation du lymphocyte B (plasmocyte/mémoire, commutation isotypique..): et obtention de la réponse anticorps secondaire |
| lymphocyte Th1 CD4 et macrophage (peu d’action des LT Th2 sur ces cellules) | - lymphocyte T spécifique d'un épitope T présenté par le macrophage via CMH2 ; lymphocyte producteur de cytokines actives sur le macrophage (IFN gamma..) |
- sécrétion de cytokines par le lymphocyte T: augmentation de l'immunité cellulaire Cf cours régulation - différenciation du lymphocyte T (mémoire..) |
| - macrophage ayant phagocyté une cible ou infecté (micro-organisme..) | - augmentation des capacités phagocytaires du macrophage (explosion respiratoire..): augmentation de l'immunité cellulaire | |
| lymphocyte T et lymphocyte NK | lymphocytes T et NK activés au cours d'un même processus (en présence de CPA qui présentent les antigènes via CMH2 aux lymphocytes T) |
- contrôle croisé, par les cytokines produites, des fonctions respectives des lymphocytes T et NK |
| mastocyte et éosinophile | mastocytes et éosinophiles activés au cours d'un même processus | - contrôle croisé, par les cytokines produites, des fonctions respectives des mastocytes et des éosinophiles |
élements d'application et de raisonnement |
| Au sens strict, la fonction de présentation immunologique ne concerne que les antigènes présentés via le CMH 2 aux lymphocytes T CD4. Toutes les cellules sont susceptibles également de présenter des antigènes issus du métabolisme cytoplasmique via le CMH 1 aux lymphocytes T CD8. Les cellules exprimant le CMH2 peuvent aussi présenter des antigènes microbiens via CD1 aux lymphocytes T gamma-delta. (fig15: cas particulier de la présentation CD1/iNKT) |
| Les capacités de présentation dépendent de nombreuses variables: - elles varient d'un individu en raison du polymorphisme du CMH; la consanguinité, en diminuant le nombre de CMH différents exprimés par un invididu, diminue les capacités immunes. - elles varient selon les modalités d'exposition à l'antigène (dose et voie d'administration), en raison des variations dans les capacités de présentation des différentes types de cellules présentatrices (par exemple les cellules impliquées dans la présentation seront différentes par voie cutanée ou digestive). - la gamme peptidique produite par un antigène donné sera différente selon la cellule présentatrice (modalités de clivage), et selon l'espèce et l'individu (allèle du CMH). |
Recemment, il a été découvert que la présentation des antigènes concerne également les lymphocytes B, dans une moindre mesure: |
| Un processus particulier de captation-présentation des antigènes a lieu dans la muqueuse digestive au niveau des plaques de Peyer (cf organes) (fig16: plaques de Peyer d'un intestin de souris) - les cellules M qui forment l'epithelium digestif sus-jacent à la plaque de Peyer sont capables de phagocyter les antigènes particulaires ou solubles présents dans le tube digestif (ces antigènes sont essentiellement des antigènes microbiens, car les antigènes alimentaires ont été presque tous dégradés en amont). - les cellules M transportent ces antigènes vers les structures lymphoïdes des plaques de Peyer, assurant une présentation des antigènes via le CMH2 aux lymphocytes, à l'origine de la réponse immune locale. |
| La coopération entre le lymphocyte B et le lymphocyte T CD4 est la mieux connue: - le lymphocyte B stimulé par l'antigène prolifère et initie la réponse anticorps primaire (IgM) - le lymphocyte B exprime également des fragments de l'antigène capté via le CMH2 - un lymphocyte T CD4 spécifique de ce même antigène reconnait l'épitope T via le CM2, s'active et produit des cytokines. Selon le profil de cytokines produit, le lymphocyte T influence la prolifération et la différenciation du lymphocyte B, et donc la réponse anticorps (ou acquérir un phénotype mémoire). - le lymphocyte B se différencie et peut produire la réponse anticorps secondaire (IgG..). Plusieurs jours sont nécessaires pour la mise en place d'une coopération efficace lors des premières expositions à l'antigène. Les différentes sous-populations CD4 ont des effets différents (cf regulation). La coopération s'effectue plus efficacement dans les follicules lymphoïdes (surtout dans les organes lymhoïdes). D'autres cellules sont souvent nécessaires pour amplifier le processus (CPA pour apporter l'antigène dans le noeud lymphatique et pour moduler la différenciation du lymphocyte T). |
| L'immunogénicité dépend à la fois de l'antigène et de l'hôte: - l'antigène immunogène doit comporter à la fois des épitopes T et B pour être reconnu par les 2 populations lymphocytaires: il s'agit très généralement d'antigènes protéiques (ou possédant une partie protéique). La production des épitopes T dépend de la qualité des mécanismes d'apprêtement (contrôlée au cours de la réponse immune). - l'hôte doit posséder à la fois des clones lymphocytaires T et B spécifiques, susceptibles d'interagir dans des mécanismes de coopération. Les mécanismes de protection du soi interviennent pour inhiber la réponse au soi et contrôler les réactions croisées entre des antigènes exogènes et des antigènes du soi: ceci explique pourquoi les protéines du soi ne sont normalement pas immunogènes. |
| La prolongation de la durée de vie des cellules mémoire est basée sur plusieurs paramètres: - fonctionnement cellulaire assurant une meilleure conservation des structures et du métabolisme cellulaire - stockage des cellules dans des structures qui fournissent un biotope adéquat (organes lymphoïdes secondaires) - augmentation du nombre de divisions possibles par une cellule (les cellules mémoire se divisent mieux et plus vite que les cellules naïves!). On considère que les cellules mémoire peuvent persister plusieurs mois (quelquefois plusieurs années) en l'absence de stimulations antigéniques. Des réactions croisées avec des antigènes similaires ou des expositions à d'infimes quantités d'antigène permettent d'entretenir l'immunité pendant des années (ex: vaccination antitétanique valable 10 ans). |
| Il est techniquement très difficile de distinguer les lymphocytes mémoire (à vie longue) des cellules naïves (à vie brève), car les conditions de culture in vitro ne préservent pas les propriétés détenues in vivo. Il existe quelques marqueurs en immunologie fondamentale (lymphocytes T mémoire: isoformes du CD45..). |
références et cours disponibles |
pour en savoir plus :
fig 7b:
Calis et al. MHC class I molecules exploit the low G+C content of pathogen genomes for enhanced presentation. Eur J Immunol (2010) vol. 40 (10) pp. 2699-709
page réalisée par le Dr Delphine Grézel, VetAgro Sup, Campus Vétérinaire de Lyon, le 28/02/13 . Merci pour les corrections, commentaires et suggestions ( delphine.grezel@vetagro-sup.fr)