 2
2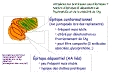 2b
2b 3 4
3 4 4b 5
4b 5 6
6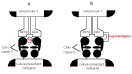
| accueil | immun1-07: Antigènes et épitopes ; réactions croisées ; soi et non soi |
<-- cours --> |
objectifs pédagogiques (*) |
|
|
- représenter sur un schéma d'un antigène les notions d'épitopes conformationnels et séquentiels | |
définitions générales - messages |
|
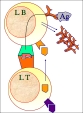
| épitope B | = épitope d'un antigène susceptible d'être reconnu par un lymphocyte B. Tous les antigènes possèdent des épitopes B. L'antigène expose en surface des épitopes B qui peuvent se fixer directement sur les anticorps ou sur les récepteurs BCR. (fig1: reconnaissance d'un antigène par les lymphocytes B et T) (cf tableau1: comparaison des épitopes T et B) |
| épitope T | = épitope d'un antigène susceptible d'être reconnu par un lymphocyte T. Tous les antigènes ne possèdent pas d'épitopes T. L'antigène, capté par une cellule, est fragmenté en épitopes T qui sont reconnus sous forme de complexe {CMH + épitope} par les récepteurs TCR. (fig1: reconnaissance d'un antigène par les lymphocytes B et T) (cf tableau1: comparaison des épitopes T et B) |
| épitope conformationnel | = épitope d'un antigène formé par la juxtaposition dans l'espace de zones de la molécule (les épitopes conformationnels peuvent être juxtaposés dans les repliements de la structure tertiaire, voire même appartenir à des molécules différentes formant un complexe multimérique). (fig2: principe des épitopes conformationnels et séquentiels) (fig2b: exemple des épitopes impliqués dans la neutralisation de la ricine) |
| épitope séquentiel | = épitope d'un antigène formé d'une succession d'éléments de l'antigène (portion de la séquence d'un antigène protéique= peptide de 5-10 AA..).(fig2: principe des épitopes conformationnels et séquentiels) |
| réaction croisée | = propriété d'un anticorps spécifique d'un antigène donné de reconnaître également un autre antigène (les 2 antigènes partagent en fait des épitopes communs). Cela peut se produire aussi avec un antisérum (mélange d'anticorps), lorsque la cible de la réponse anticorps possède des antigènes communs avec d'autres structure/organismes (fig3: réactions croisées à l'échelle d'un antigène ou fig4: d'un micro-organisme) . |
| La majorité des réactions croisées est due à des homologies entre les antigènes (=portions de séquence communes..) ou à des parentés entre les organismes ou micro-organismes (bactéries d'un même genre ou d'une même espèce..) : ceux-ci présentent alors une proportion non négligeable d'antigènes communs ou peu différents.. Généralement les réactions croisées sont incomplètes (l'affinité pour le 2ème antigène est < à l'affinité pour le 1er), et ne concernent qu'une partie des antigènes d'un micro-organisme (antigènes communs). Les réactions croisées ont un rôle biologique important (par exemple: réponse à un Ag2 supérieure à celle attendue en raison d’une première stimulation par un Ag1 apparenté). |
|
Les réactions croisées
permettent d'obtenir rapidemment une réponse immune vis à vis d'un agent infectieux, si on a déjà rencontré des antigènes présentant des épitopes identiques auparavant (vaccin, micro-organisme commensal ou infectieux apparenté). Elles permettent aussi de prolonger l'immunité contre un agent infectieux au contact d'autres agents. |
|
| sérotype | = sous-groupe identifié au sein d'une espèce en fonction de ses propriétés antigéniques, à l'aide d'un anticorps ou d'un antisérum de référence.Les variations intraspécifiques des antigènes sont fréquentes chez les µorganismes et sont à l’origine de souches qui possèdent une distribution géographique ou une virulence particulière (espèce-cible, gravité...). Une souche particulière au sein d'une espèce bactérienne peut être identifiée par son sérotype (idem sérotype viral). D'où l'intérêt d'identifier le sérotype au cours du diagnostic d'une infection, pour disposer de données sur sa gravité ou son épidémiologie. Les groupes sanguins et tissulaires d'un individu peuvent également être identifiés à l'aide d'anticorps de référence (sérotypes). |
| antigénicité | = aptitude d'un antigène à être reconnu par une réponse immune (jusqu'à ce que l'antigène soit entièrement éliminé de l'organisme). |
| haptène | = molécule qui peut réagir spécifiquement avec un anticorps, mais qui ne peut pas déclencher la synthèse de cet anticorps chez un animal : ce n’est pas un antigène à proprement parler (et encore moins un immunogène): petite taille, instabilité.. En revanche, si un haptène établit une liaison covalente avec une protéine porteuse, l'ensemble induit alors une réponse anticorps, de type secondaire (IgG..). Cette fixation peut être obtenue au laboratoire, artificiellement (fabrication de réactifs contre de petites molécules comme des hormones en les fixant à une protéine porteuse), ou arriver spontanément dans certaines maladies de l’immunité contre des xénobiotiques (protéine porteuse = protéine normale de l’organisme qui établit «malencontreusement» des liaisons avec le xénobiotique: ex allergie pénicilline) (fig5: haptène) |
| super-antigène | = molécule capable de provoquer une activation polyclonale des lymphocytes T en se fixant sur le récepteur (TCR) sans tenir compte de sa spécificité, car elle se fixe sur des domaines constants des chaines du TCR (fig6: comparaison antigène et super-antigène). On connaît quelques dizaine de superantigènes : ils sont impliqués dans la toxicité de certains poisons et la virulence de certains micro-organismes car ils désorganisent la réponse immune (activation anarchique de nombreux clones lymphocytaires T). |
| soi et non-soi (cf immun1-17) | = concepts utilisés en immunologie pour expliquer comment la réponse immune arrive à distinguer les antigènes appartenant en propre à l'individu (=soi) et les antigènes exogènes (non-soi). Les antigènes exogènes sont extrêmement nombreux et divers: antigènes microbiens et parasitaires, xénobiotiques (médicaments..) et molécules étrangères qui réussissent à passer les barrières naturelles (corps étrangers, antigènes alimentaires ou inhalés..). Normalement, des mécanismes de régulation et d'éducation empêchent les antigènes du soi de déclencher une réponse immune (alors qu'ils peuvent être immunogènes et provoquer des réactions de rejet si ils sont administrés à une espèce ou à un individu différent). |
schémas et figures |
|
tableaux |
tableau 2: exemples de biodiversité liées aux variations antigéniques inter et intra-spécifiques |
micro-organismes, fungi et parasites ; invertébrés | variations inter-spécifiques chez vertébrés | variations intra-spécifiques chez les vertébrés |
| antigènes caractéristiques d'une espèce (nombreux épitopes reconnus) | - nombreuses molécules caractéristiques du monde microbien, des protozoaires et des fungi: paroi bactérienne, endotoxines et autres molécules typiquement microbiennes (sucres complexes..), protéines typiquement microbiennes (toxines, certaines enzymes..) (fig4b: principaux antigènes des bactéries) - nombreuses molécules caractéristiques des invertébrés (hemocyanine, chitine, soie, GFP..).. |
il n'existe que quelques exemples d'antigènes caractéristiques d'espèce au sein des vertébrés: protéines des venins.. | Il existe quelques protéines codées par le chromosome Y (expression exclusive chez le mâle). |
| Du fait de la très grande plasticité du génome microbien, les molécules (protéines, endotoxines..) présentent des variations importantes d'une espèce à l'autre, voire même entre souches au sein d'une même espèce (molécules distinctes mais susceptibles de produire des réactions croisées). ex: des centaines de sérotypes connus pour les antigènes K, F, O d'Escherichia coli (dont certains identifient des souches pathogènes, en particulier pour l'homme, le veau, le porc..). |
|||
| antigènes présents sous des formes similaires mais légèrement différentes (variations concernant quelques épitopes) | quelques protéines, en particulier des enzymes, sont conservées dans l'ensemble du règne animal avec de faibles variations inter-spécifiques | La majorité des protéines existent sous des formes similaires chez tous les vertébrés: les homologies sont plus ou moins grandes en fonction de la proximité phylogénétique (l'antigénicité est due à quelques épitopes dissemblables d'une espèce à l'autre ) | Quelques protéines subissent des variations intra-spécifiques liées à des variations alléliques, ou à l'expression différenciée selon les individus de librairies génétiques complexes (groupes tissulaires et groupes sanguins, allotypes..) |
| ex: l'albumine du chien présente 76% d'homologie avec l'albumine bovine, 74% avec l'albumine de lapin et 47% avec l'albumine de poule (ovalbumine). | ex: il existe 23 variants de la protéine HLA-A chez l'homme (l'allèle le plus répandu est partagé par 5% de la population) | ||
| antigènes identiques | La majorité des molécules organiques simples sont quasi-identiques dans l'ensemble du règne animal (sucres, acides gras, neuromédiateurs et hormones non protéiques..). | ||
élements d'application et de raisonnement |
| L'antigénicité dépend du nombre d'épitopes et de la stabilité de l'antigène. Seuls certains critères sont requis pour être un antigène : - taille > 1kDa (équivalent d'une dizaine de cycles aromatiques). Les petites molécules (eau, mineraux, hormones stéroïdes, vitamines..) ne sont pas antigèniques. En revanche, certaines petites molécules (pas toutes! : certains métaux comme le nickel, certains médicaments comme la pénicilline..) peuvent s'associer à des protéines cellulaires ou plasmatiques, les dénaturant au point qu'elles deviennent antigéniques. - stabilité et biodisponibilité. Les molécules facilement détruites dans les tissus (par les enzymes, le pH..) ne sont pas antigéniques car elles ne persistent pas assez longtemps pour donner lieu à une réponse. De plus, la capacité de la molécule à diffuser dans différents tissus et liquides biologiques (hydrophilie..) conditionne sa rencontre avec des lymphocytes, et donc la possibilité d'obtenir sa reconnaissance spécifique. |
| De très nombreuses molécules sont des antigènes, y compris nos propres constituants (groupes sanguins..). Pour simplifier, on peut considèrer qu'un virus correspond à peu près à 20-100 antigènes différents, une bactérie à 200-2000 antigènes différents et un organisme eucaryote (protozoaire, invertébré ou vertébré) à plus de 5000 antigènes qui peuvent donner lieu à une reconnaissance spécifique (mais une minorité est immunogène) (cf tableau2: biodiversité). |
| Les sérotypes microbiens correspondent à des variants antigéniques au sein d'une espèce microbienne (souches), de même que les biotypes et les génotypes correspondent respectivement à des variants métaboliques ou génétiques. Comme beaucoup d'antigènes jouent un rôle dans la virulence (toxines, facteurs d'adhésion, enzymes..), il est fréquent de distinguer des sérotypes "pathogènes" et des sérotypes "moins pathogènes" (par exemple E.coli O157 responsable d'infection grave chez l'homme, alors que la majorité des E.coli ne sont pas pathogènes, différents sérotypes de Leptospires..). Pour identifier des sérotypes, le laboratoire doit disposer d'anticorps de référence produits contre des souches de référence cultivées au laboratoire (la majorité sont maintenant des anticorps monoclonaux). |
| Les réactions croisées entre micro-organismes ont un rôle biologique: l'individu qui a élaboré une réponse immune contre un micro-organisme opportuniste est plus apte à se défendre contre un micro-organisme pathogène apparenté (réactions croisées contre une partie des antigènes communs). En revanche, il faut savoir interpréter judicieusement les tests de diagnostic en fonction des réactions croisées possibles (différentes astuces techniques sont utilisées pour se débarasser des réactions croisées, de façon à augmenter la spécificité des tests). |
| Les réactions croisées sont des phénomènes fréquents, mais la grande majorité de ces réactions ont une affinité faible ou ne concernent qu'une partie des épitopes (d'où la grande spécificité des réponses immunes). |
| Certaines réactions croisées entre des antigènes étrangers et des protéines de l'organisme peuvent occasionner des troubles immunopathologiques sévères (Streptococus pyogenes/rhumatisme articulaire aigü, Maladie de Chagas/cardiomyopathie..) |
| Il peut paraitre étonnant qu'une protéine du soi puisse induire une réponse anticorps lorsqu'un haptène y est fixé : on considère que l'haptène apporte l'épitope B et la protéine l'épiptope T nécessaires à la mise en place d'une forte réponse anticorps. L'ensemble est immunogène même si la protéine porteuse appartient au soi, car la protéine peut-être dénaturée par la fixation de l'haptène (d'où la création d'épitopes T inhabituels), ou qu'un contexte activateur/inflammatoire supprime l'anergie naturelle vis-à-vis des épitopes T de cette protéine (dans la mesure où tous les clones T auto-réactifs ne sont pas éliminés par l'éducation thymique). |
- quels sont les types d'épitopes d'un haptène ? d'un superantigène?
- envisager différents moyens de réduire les réactions croisées dans un test de diagnostic.
références et cours disponibles |
pour en savoir plus :
- identification des épitopes (exemple de l'osteocalcine bovine): http://bio.takara.co.jp/BIO_EN/catalog_d.asp?C_ID=C0501
page réalisée par le Dr Delphine Grézel, VetAgro Sup, Campus Vétérinaire de Lyon, le 1/02/12 . Merci pour les corrections, commentaires et suggestions ( delphine.grezel@vetagro-sup.fr)